terça-feira, 20 de novembro de 2012
domingo, 12 de agosto de 2012
GENÉTICA
Genética
Desde os tempos mais remotos o homem
tomou consciência da importância do macho e da fêmea na geração de seres
da mesma espécie, e que características como altura, cor da pele etc.
eram transmitidas dos pais para os descendentes. Assim, com certeza, uma
cadela quando cruzar com um cão, irá originar um filhote com
características de um cão e nunca de um gato. Mas por quê?
Mendel, o iniciador da genética
Gregor Mendel nasceu em 1822, em
Heinzendorf, na Áustria. Era filho de pequenos fazendeiros e, apesar de
bom aluno, teve de superar dificuldades financeiras para conseguir
estudar. Em 1843, ingressou como noviço no mosteiro de agostiniano da
cidade de Brünn, hoje Brno, na atual República Tcheca.
 Após ter sido
ordenado monge, em 1847, Mendel ingressou na Universidade de Viena, onde
estudou matemática e ciências por dois anos. Ele queria ser professor
de ciências naturais, mas foi mal sucedido nos exames. Após ter sido
ordenado monge, em 1847, Mendel ingressou na Universidade de Viena, onde
estudou matemática e ciências por dois anos. Ele queria ser professor
de ciências naturais, mas foi mal sucedido nos exames.
De volta a Brünn, onde passou o
resto da vida. Mendel continuou interessado em ciências. Fez estudos
meteorológicos, estudou a vida das abelhas e cultivou plantas, tendo
produzido novas variedades de maças e peras. Entre 1856 e 1865, realizou
uma série de experimentos com ervilhas, com o objetivo de entender como
as características hereditárias eram transmitidas de pais para filhos.
Em 8 de março de 1865, Mendel
apresentou um trabalho à Sociedade de História Natural de Brünn, no qual
enunciava as suas leis de hereditariedade, deduzidas das experiências
com as ervilhas. Publicado em 1866, com data de 1865, esse trabalho
permaneu praticamente desconhecido do mundo científico até o início do
século XX. Pelo que se sabe, poucos leram a publicação, e os que leram
não conseguiram compreender sua enorme importância para a Biologia. As
leis de Mendel foram redescobertas apenas em 1900, por três
pesquisadores que trabalhavam independentemente.
|
Mendel morreu em Brünn, em 1884. Os
últimos anos de sua vida foram amargos e cheios de desapontamento. Os
trabalhos administrativos do mosteiro o impediam de se dedicar
exclusivamente à ciência, e o monge se sentia frustrado por não ter
obtido qualquer reconhecimento público pela sua importante descoberta.
Hoje Mendel é tido como uma das figuras mais importantes no mundo
científico, sendo considerado o “pai” da Genética. No
mosteiro onde viveu existe um monumento em sua homenagem, e os jardins
onde foram realizados os célebres experimentos com ervilhas até hoje são
conservados.
Os experimentos de Mendel
A escolha da planta
A ervilha é uma planta herbácea
leguminosa que pertence ao mesmo grupo do feijão e da soja. Na
reprodução, surgem vagens contendo sementes, as ervilhas. Sua escolha
como material de experiência não foi casual: uma planta fácil de
cultivar, de ciclo reprodutivo curto e que produz muitas sementes. Desde
os tempos de Mendel existiam muitas variedades disponíveis, dotadas de
características de fácil comparação. Por exemplo, a variedade que flores
púrpuras podia ser comparada com a que produzia flores brancas; a que
produzia sementes lisas poderia ser comparada cm a que produzia sementes
rugosas, e assim por diante. Outra vantagem dessas plantas é que estame
e pistilo, os componentes envolvidos na reprodução sexuada do vegetal,
ficam encerrados no interior da mesma flor, protegidas pelas pétalas.
Isso favorece a autopolinização e, por extensão, a autofecundação,
formando descendentes com as mesmas características das plantas
genitoras.
 A partir da autopolinização, Mendel
produziu e separou diversas linhagens puras de ervilhas para as
características que ele pretendia estudar. Por exemplo, para cor de
flor, plantas de flores de cor de púrpura sempre produziam como
descendentes plantas de flores púrpuras, o mesmo ocorrendo com o
cruzamento de plantas cujas flores eram brancas. Mendel estudou sete
características nas plantas de ervilhas: cor da flor, posição da flor no
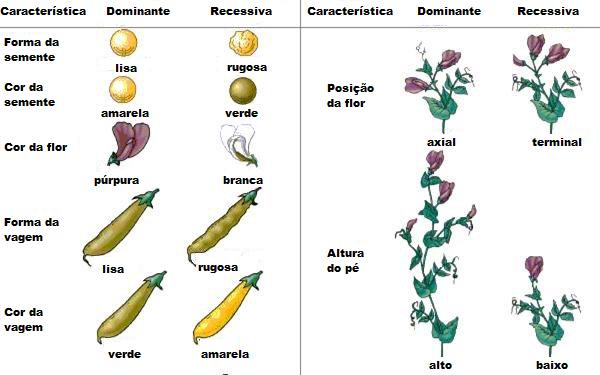
caule, cor da semente, aspecto externo da semente, forma da vagem, cor
da vagem e altura da planta.
A partir da autopolinização, Mendel
produziu e separou diversas linhagens puras de ervilhas para as
características que ele pretendia estudar. Por exemplo, para cor de
flor, plantas de flores de cor de púrpura sempre produziam como
descendentes plantas de flores púrpuras, o mesmo ocorrendo com o
cruzamento de plantas cujas flores eram brancas. Mendel estudou sete
características nas plantas de ervilhas: cor da flor, posição da flor no
caule, cor da semente, aspecto externo da semente, forma da vagem, cor
da vagem e altura da planta.
Os cruzamentos
Depois de obter linhagens puras,
Mendel efetuou um cruzamento diferente. Cortou os estames de uma flor
proveniente de semente verde e depois depositou, nos estigmas dessa
flor, pólen de uma planta proveniente de semente amarela. Efetuou,
então, artificialmente, uma polinização cruzada: pólen
de uma planta que produzia apenas semente amarela foi depositado no
estigma de outra planta que só produzia semente verde, ou seja, cruzou
duas plantas puras entre si. Essas duas plantas foram consideradas como a
geração parental (P), isto é, a dos genitores.
 |
Após repetir o mesmo procedimento
diversas vezes, Mendel verificou que todas as sementes originadas
desses cruzamentos eram amarelas – a cor verde havia aparentemente
“desaparecido” nos descendentes híbridos (resultantes do cruzamento das
plantas), que Mendel chamou de F1 (primeira geração filial). Concluiu, então, que a cor amarela “dominava” a cor verde. Chamou o caráter cor amarela da semente de dominante e o verde de recessivo.
A seguir, Mendel fez germinar as sementes obtidas em F1
até surgirem as plantas e as flores. Deixou que se autofertilizassem e
aí houve a surpresa: a cor verde das sementes reapareceu na F2
(segunda geração filial), só eu em proporção menor que as de cor
amarela: surgiram 6.022 sementes amarelas para 2.001 verdes, o que
conduzia a proporção 3:1. Concluiu que na verdade, a cor verde das sementes não havia “desaparecido” nas sementes da geração F1.
O que ocorreu é que ela não tinha se manifestado, uma vez que, sendo
uma caráter recessivo, era apenas “dominado” (nas palavras de Mendel)
pela cor amarela. Mendel concluiu que a cor das sementes era determinada
por dois fatores, cada um determinando o surgimento de uma cor, amarela
ou verde.
|
Era necessário definir uma simbologia
para representar esses fatores: escolheu a inicial do caráter recessivo.
Assim, a letra v (inicial de verde), minúscula, simbolizava o fator
recessivo. Assim, a letra v (inicial de verde), minúscula,
simbolizava o fator recessivo – para cor verse – e a letra V, maiúscula,
o fator dominante – para cor amarela.
VV
|
vv
|
Vv
|
Semente amarela pura
|
Semente verde pura
|
Semente amarela híbrida
|
Persistia, porém, uma dúvida: Como explicar o desaparecimento da cor verde na geração F1 e o seu reaparecimento na geração F2?
A resposta surgiu a partir do
conhecimento de que cada um dos fatores se separava durante a formação
das células reprodutoras, os gametas. Dessa forma, podemos entender como
o material hereditário passa de uma geração para a outra. Acompanhe nos
esquemas abaixo os procedimentos adorados por Mendel com relação ao
caráter cor da semente em ervilhas.

Resultado: em F2,
para cada três sementes amarelas, Mendel obteve uma semente de cor
verde. Repetindo o procedimento para outras seis características
estudadas nas plantas de ervilha, sempre eram obtidos os mesmos
resultados em F2, ou seja a proporção de três expressões dominantes para uma recessiva.
Leis de Mendel
1ª Lei de Mendel: Lei da Segregação dos Fatores
A comprovação da hipótese de dominância
e recessividade nos vários experimentos efetuados por Mendel levou,
mais tarde à formulação da sua 1º lei: “Cada característica é
determinada por dois fatores que se separam na formação dos gametas,
onde ocorrem em dose simples”, isto é, para cada gameta masculino ou feminino encaminha-se apenas um fator.
Mendel não tinha idéia da constituição desses fatores, nem onde se localizavam.
As bases celulares da segregação
A redescoberta dos trabalhos de Mendel, em 1900, trouxe a questão: onde estão os fatores hereditários e como eles se segregam?
 |
Em 1902, enquanto estudava a
formação dos gametas em gafanhotos, o pesquisador norte americano Walter
S. Sutton notou surpreendente semelhança entre o comportamento dos
cromossomos homólogos, que se separavam durante a meiose, e os fatores
imaginados por Mendel. Sutton lançou a hipótese de que os pares de
fatores hereditários estavam localizados em pares de cromossomos
homólogos, de tal maneira que a separação dos homólogos levava à
segregação dos fatores.
Hoje sabemos que os fatores a que Mendel se referiu são os genes (do grego genos,
originar, provir), e que realmente estão localizados nos cromossomos,
como Sutton havia proposto. As diferentes formas sob as quais um gene
pode se apresentar são denominadas alelos. A cor amarela e a cor verde
da semente de ervilha, por exemplo, são determinadas por dois alelos,
isto é, duas diferentes formas do gene para cor da semente.
|
Exemplo da primeira lei de Mendel em um animal
Vamos estudar um exemplo da aplicação
da primeira lei de Mendel em um animal, aproveitando para aplicar a
terminologia modernamente usada em Genética. A característica que
escolhemos foi a cor da pelagem de cobaias, que pode ser preta ou
branca. De acordo com uma convenção largamente aceita, representaremos
por B o alelo dominante, que condiciona a cor preta, e por b o alelo
recessivo, que condiciona a cor branca.
Uma técnica simples de combinar os gametas produzidos pelos indivíduos de F1 para obter a constituição genética dos indivíduos de F2
é a montagem do quadrado de Punnet. Este consiste em um quadro, com
número de fileiras e de colunas que correspondem respectivamente, aos
tipos de gametas masculinos e femininos formados no cruzamento. O
quadrado de Punnet para o cruzamento de cobaias heterozigotas é:
B
Gametas paternos
b
|
Gametas maternos
B b
|
|
BB
Preto
|
Bb
Preto
|
|
Bb
Preto
|
bb
Branco
|
|
Os conceitos de fenótipo e genótipo
Dois conceitos importantes para o desenvolvimento da genética, no começo do século XX, foram os de fenótipo e genótipo, criados pelo pesquisador dinamarquês Wilhelm L. Johannsen (1857 – 1912).
Fenótipo
O termo “fenótipo” (do grego pheno, evidente, brilhante, e typos,
característico) é empregado para designar as características
apresentadas por um indivíduo, sejam elas morfológicas, fisiológicas e
comportamentais. Também fazem parte do fenótipo características
microscópicas e de natureza bioquímica, que necessitam de testes
especiais para a sua identificação.
Entre as características fenotípicas visíveis,
podemos citar a cor de uma flor, a cor dos olhos de uma pessoa, a
textura do cabelo, a cor do pelo de um animal, etc. Já o tipo sanguíneo e
a sequência de aminoácidos de uma proteína são características
fenotípicas revelada apenas mediante testes especiais.

O fenótipo de um indivíduo sofre transformações com o
passar do tempo. Por exemplo, à medida que envelhecemos o nosso corpo
se modifica. Fatores ambientais também podem alterar o fenótipo: se
ficarmos expostos à luz do sol, nossa pele escurecerá.
Genótipo
O termo “genótipo” (do grego genos, originar, provir, e typos, característica) refere-se à constituição genética do indivíduo,
ou seja, aos genes que ele possui. Estamos nos referindo ao genótipo
quando dizemos, por exemplo, que uma planta de ervilha é homozigota
dominante (VV) ou heterozigota (Vv) em relação à cor da semente.
Fenótipo: genótipo e ambiente em interação
O fenótipo resulta da interação do genótipo com o
ambiente. Consideremos, por exemplo, duas pessoas que tenham os mesmos
tipos de alelos para pigmentação da pele; se uma delas toma sol com mais
frequência que a outra, suas tonalidades de pele, fenótipo, são
diferentes.
Um exemplo interessante de interação entre genótipo e
ambiente na produção do fenótipo é a reação dos coelhos da raça
himalaia à temperatura. Em temperaturas baixas, os pelos crescem pretos
e, em temperaturas altas, crescem brancos. A pelagem normal desses
coelhos é branca, menos nas extremidades do corpo (focinho, orelha, rabo
e patas), que, por perderem mais calor e apresentarem temperatura mais
baixa, desenvolvem pelagem preta.
Determinando o genótipo
Enquanto que o fenótipo de um indivíduo pode ser
observado diretamente, mesmo que seja através de instrumentos, o
genótipo tem que ser inferido através da observação do fenótipo, da
análise de seus pais, filhos e de outros parentes ou ainda pelo
sequenciamento do genoma do indivíduo, ou seja, leitura do que está nos
genes. A técnica do sequenciamento, não é amplamente utilizada, devido
ao seu alto custo e pela necessidade de aparelhagem especializada. Por
esse motivo a observação do fenótipo e análise dos parentes ainda é o
recurso mais utilizado para se conhecer o genótipo.
Quando um indivíduo apresenta o fenótipo condicionado
pelo alelo recessivo, conclui-se que ele é homozigoto quanto ao alelo
em questão. Por exemplo, uma semente de ervilha verde é sempre
homozigota vv. Já um indivíduo que apresenta o fenótipo condicionado
pelo alelo dominante poderá ser homozigoto ou heterozigoto. Uma semente
de ervilha amarela, por exemplo, pode ter genótipo VV ou Vv. Nesse caso,
o genótipo do indivíduo só poderá ser determinado pela análise de seus
pais e de seus descendentes.
Caso o indivíduo com fenótipo dominante seja filho
de pai com fenótipo recessivo, ele certamente será heterozigoto, pois
herdou do pai um alelo recessivo. Entretanto, se ambos os pais têm
fenótipo dominante, nada se pode afirmar. Será necessário analisar a
descendência do indivíduo em estudo: se algum filho exibir o fenótipo
recessivo, isso indica que ele é heterozigoto.
 |
Cruzamento-teste
Este cruzamento é feito com um indivíduo
homozigótico recessivo para o fator que se pretende estudar, que
facilmente se identifica pelo seu fenótipo e um outro de genótipo
conhecido ou não. Por exemplo, se cruzarmos um macho desconhecido com
uma fêmea recessiva podemos determinar se o macho é portador daquele
caráter recessivo ou se é puro. Caso este seja puro todos os filhos
serão como ele, se for portador 25% serão brancos, etc. Esta explicação é
muito básica, pois geralmente é preciso um pouco mais do que este único
cruzamento.
A limitação destes cruzamentos está no fato de
não permitirem identificar portadores de alelos múltiplos para a mesma
característica, ou seja, podem existir em alguns casos mais do que dois
alelos para o mesmo gene e o efeito da sua combinação variar. Além
disso, podemos estar cruzando um fator para o qual o macho ou fêmea
teste não são portadores, mas sim de outros alelos.
|
Construindo um heredograma
No caso da espécie humana, em que não se pode
realizar experiências com cruzamentos dirigidos, a determinação do
padrão de herança das características depende de um levantamento do
histórico das famílias em que certas características aparecem. Isso
permite ao geneticista saber se uma dada característica é ou não
hereditária e de que modo ela é herdada. Esse levantamento é feito na
forma de uma representação gráfica denominada heredograma (do latim heredium, herança), também conhecida como genealogia ou árvore genealógica.
Construir um heredograma consiste em representar,
usando símbolos, as relações de parentesco entre os indivíduos de uma
família. Cada indivíduo é representado por um símbolo que indica as suas
características particulares e sua relação de parentesco com os demais.
Indivíduos do sexo masculino são representados por
um quadrado, e os do sexo feminino, por um círculo. O casamento, no
sentido biológico de procriação, é indicado por um traço horizontal que
une os dois membros do casal. Os filhos de um casamento são
representados por traços verticais unidos ao traço horizontal do casal.
Os principais símbolos são os seguintes:

A montagem de um heredograma obedece a algumas regras:
1ª) Em cada casal, o homem deve ser colocado à esquerda, e a mulher à direita, sempre que for possível.
2ª) Os filhos devem ser colocados em ordem de nascimento, da esquerda para a direita.
3ª) Cada geração que se sucede é
indicada por algarismos romanos (I, II, III, etc.). Dentro de cada
geração, os indivíduos são indicados por algarismos arábicos, da
esquerda para a direita. Outra possibilidade é se indicar todos os
indivíduos de um heredograma por algarismos arábicos, começando-se pelo
primeiro da esquerda, da primeira geração.
Interpretação dos Heredogramas
A análise dos heredogramas pode permitir se
determinar o padrão de herança de uma certa característica (se é
autossômica, se é dominante ou recessiva, etc.). Permite, ainda,
descobrir o genótipo das pessoas envolvidas, se não de todas, pelo menos
de parte delas. Quando um dos membros de uma genealogia manifesta um
fenótipo dominante, e não conseguimos determinar se ele é homozigoto
dominante ou heterozigoto, habitualmente o seu genótipo é indicado como A_, B_ou C_, por exemplo.
A primeira informação que se procura obter, na análise de um heredograma, é se o caráter em questão é condicionado por um gene dominante ou recessivo.
Para isso, devemos procurar, no heredograma, casais que são
fenotipicamente iguais e tiveram um ou mais filhos diferentes deles. Se a
característica permaneceu oculta no casal, e se manifestou no filho, só
pode ser determinada por um gene recessivo. Pais fenotipicamente
iguais, com um filho diferente deles, indicam que o caráter presente no
filho é recessivo!
Uma vez que se descobriu qual é o gene dominante e
qual é o recessivo, vamos agora localizar os homozigotos recessivos,
porque todos eles manifestam o caráter recessivo. Depois disso, podemos
começar a descobrir os genótipos das outras pessoas. Devemos nos lembrar
de duas coisas:
1ª) Em um par de genes alelos, um veio do pai e o
outro veio da mãe. Se um indivíduo é homozigoto recessivo, ele deve ter
recebido um gene recessivo de cada ancestral.
2ª) Se um indivíduo é homozigoto recessivo, ele envia
o gene recessivo para todos os seus filhos. Dessa forma, como em um
“quebra-cabeças”, os outros genótipos vão sendo descobertos. Todos os
genótipos devem ser indicados, mesmo que na sua forma parcial (A_, por exemplo).
Exemplo:

Em uma árvore desse tipo, as mulheres são
representadas por círculos e os homens por quadrados. Os casamentos são
indicados por linhas horizontais ligando um círculo a um quadrado. Os
algarismos romanos I, II, III à esquerda da genealogia representam as
gerações. Estão representadas três gerações. Na primeira há uma mulher e
um homem casados, na segunda, quatro pessoas, sendo três do sexo
feminino e uma do masculino. Os indivíduos presos a uma linha horizontal
por traços verticais constituem uma irmandade. Na segunda geração
observa-se o casamento de uma mulher com um homem de uma irmandade de
três pessoas.
Dominância incompleta ou Co-dominância
Nem todas as características são herdadas como a cor
da semente da ervilha, em que o gene para a cor amarela domina sobre o
gene para cor verde. Muito frequentemente a combinação dos genes alelos
diferentes produz um fenótipo intermediário. Essa situação ilustra a chamada dominância incompleta ou parcial.
Um exemplo desse tipo de herança é a cor das flores maravilha. Elas
podem ser vermelhas, brancas ou rosas. Plantas que produzem flores
cor-de-rosa são heterozigotas, enquanto os outros dois fenótipos são
devidos à condição homozigota. Supondo que o gene V determine a cor vermelha e o gene B, cor branca, teríamos:
VV = flor vermelha
BB = flor branca
VB = flor cor-de-rosa
Apesar de anteriormente usarmos letras maiúsculas
para indicar, respectivamente, os genes dominantes e recessivos, quando
se trata de dominância incompleta muitos autores preferem utilizar
apenas diferentes letras maiúsculas.
Fazendo o cruzamento de uma planta de maravilha que
produz flores vermelhas com outra que produz flores brancas e analisando
os resultados fenotípicos da geração F1e F2, teríamos:

Agora analizando os resulados genotípicos da geração F1e F2, teríamos:
P:
|
Flor Branca
B B
|
|
V
Flor Vermelha
V
|
BV
cor-de-rosa
|
BV
cor-de-rosa
|
VB
cor-de-rosa
|
VB
cor-de-rosa
|
|
F1 = 100% VB (flores cor-de-rosa)
Cruzando, agora, duas plantas heterozigotas (flores cor-de-rosa), teríamos:
F1
|
Flor cor-de-rosa
V B
|
|
V
Flor cor-de-rosa
B
|
VV
Vermelha
|
BV
cor-de-rosa
|
VB
cor-de-rosa
|
BB
Branca
|
|
F2 = Genótipos: 1/4 VV, 1/2 VB, 1/4 BB.
Fenótipo: 1/4 plantas com flores vermelhas
1/2 plantas com flores cor-de-rosa
1/4 plantas com flores brancas
Alelos letais: Os genes que matam
As mutações que ocorrem nos seres vivos são
totalmente aleatórias e, às vezes, surgem variedades genéticas que podem
levar a morte do portador antes do nascimento ou, caso ele sobreviva,
antes de atingir a maturidade sexual. Esses genes que conduzem à morte
do portador, são conhecidos como alelos letais. Por exemplo, em uma
espécie de planta existe o gene C, dominante, responsável pela coloração verde das folhas. O alelo recessivo c, condiciona a ausência de coloração nas folhas, portanto o homozigoto recessivo cc
morre ainda na fase jovem da planta, pois esta precisa do pigmento
verde para produzir energia através da fotossíntese. O heterozigoto é
uma planta saudável, mas não tão eficiente na captação de energia solar,
pela coloração verde clara em suas folhas. Assim, se cruzarmos duas
plantas heterozigotas, de folhas verdes claras, resultará na proporção 2:1 fenótipos entre os descendentes, ao invés da proporção de 3:1
que seria esperada se fosse um caso clássico de monoibridismo
(cruzamento entre dois indivíduos heterozigotos para um único gene). No
caso das plantas o homozigoto recessivo morre logo após germinar, o que
conduz a proporção 2:1.
P
|
Planta com folhas verde claras
C c
|
|
C
Planta com folhas verde claras
c
|
CC
Verde escuro
|
Cc
Verde clara
|
Cc
Verde clara
|
cc
Inviável
|
|
F1 = Fenótipo: 2/3 Verde clara
1/3 Verde escura
Genótipo: 2/3 Cc
1/3 CC
Esse curioso caso de genes letais foi descoberto em
1904 pelo geneticista francês Cuénot, que estranhava o fato de a
proporção de 3:1 não ser obedecida. Logo, concluiu se tratar de uma caso
de gene recessivo que atuava como letal quando em dose dupla.
No homem, alguns genes letais provocam a morte do feto. É o caso dos genes para acondroplasia,
por exemplo. Trata-se de uma anomalia provocada por gene dominante que,
em dose dupla, acarreta a morte do feto, mas em dose simples ocasiona
um tipo de nanismo, entre outras alterações.
Há genes letais no homem, que se manifestam depois do nascimento, alguns na infância e outros na idade adulta. Na infância, por exemplo, temos os causadores da fibrose cística e da distrofia muscular de Duchenne
(anomalia que acarreta a degeneração da bainha de mielina nos nervos).
Dentre os que se expressam tardiamente na vida do portador, estão os
causadores da doença de Huntington,
em que há a deterioração do tecido nervoso, com perde de células
principalmente em uma parte do cérebro, acarretando perda de memória,
movimentos involuntários e desequilíbrio emocional.
Como os genes se manifestam
 |
Vimos que, em alguns
casos, os genes se manifestam com fenótipos bem distintos. Por exemplo,
os genes para a cor das sementes em ervilhas manifestam-se com fenótipos
bem definidos, sendo encontradas sementes amarelas ou verdes. A essa
manifestação gênica bem determinada chamamos de variação gênica descontínua, pois não há fenótipos intermediários.
Há herança de características, no entanto,
cuja manifestação do gene (também chamada de expressividade) não
determina fenótipos tão definidos, mas sim uma gradação de fenótipos. A
essa gradação da expressividade do gene, variando desde um fenótipo que
mostra leve expressão da característica até sua expressão total,
chamamos de norma de reação ou expressividade variável.
Por exemplo, os portadores dos genes para braquidactilia (dedos curto)
podem apresentar fenótipos variando de dedos levemente mais curtos até a
total falta deles.
Alguns genes sempre que estão presentes se manifestam, dizemos que são altamente penetrantes. Outros possuem uma penetrância incompleta, ou seja, apenas uma parcela dos portadores do genótipo apresenta o fenótipo correspondente.
Observe que o conceito de penetrância está
relacionado à expressividade do gene em um conjunto de indivíduos, sendo
apresentado em termos percentuais. Assim, por exemplo, podemos falar
que a penetrância para o gene para a doença de Huntington é de 100%, o
que quer dizer que 100% dos portadores desse gene apresentam (expressam)
o fenótipo correspondente.
|
|

Diferentes graus de braquidactilia pela expressão variável do genótipo.
|
Noções de probabilidade aplicadas à genética
Acredita-se que um dos motivos para as idéias de
Mendel permanecerem incompreendidas durante mais de 3 décadas foi o
raciocínio matemático que continham. Mendel partiu do princípio que a
formação dos gametas seguia as leis da probabilidade, no tocante a
distribuição dos fatores.
Princípios básicos de probabilidade
Probabilidade é a chance que um evento tem de
ocorrer, entre dois ou mais eventos possíveis. Por exemplo, ao lançarmos
uma moeda, qual a chance dela cair com a face “cara” voltada para cima?
E em um baralho de 52 cartas, qual a chance de ser sorteada uma carta
do naipe ouros?
 |
Eventos aleatórios
Eventos como obter “cara” ao lançar uma moeda,
sortear um “ás” de ouros do baralho, ou obter “face 6” ao jogar um dado
são denominados eventos aleatórios (do latim alea, sorte) porque cada um deles tem a mesma chance de ocorrer em relação a seus respectivos eventos alternativos.
Veja a seguir as probabilidades de ocorrência
de alguns eventos aleatórios. Tente explicar por que cada um deles
ocorre com a probabilidade indicada.
|
- A probabilidade de sortear uma carta de espadas de um baralho de 52 cartas é de ¼
- A probabilidade de sortear um rei qualquer de um baralho de 52 cartas é de 1/13.
- A probabilidade de sortear o rei de espadas de um baralho de 52 cartas é de 1/52.
A formação de um determinado tipo de gameta, com um
outro alelo de um par de genes, também é um evento aleatório. Um
indivíduo heterozigoto Aa tem a mesma probabilidade de formar gametas portadores do alelo A do que de formar gametas com o alelo a (1/2 A: 1/2 a).
Eventos independentes
Quando a ocorrência de um evento não afeta a
probabilidade de ocorrência de um outro, fala-se em eventos
independentes. Por exemplo, ao lançar várias moedas ao mesmo tempo, ou
uma mesma moeda várias vezes consecutivas, um resultado não interfere
nos outros. Por isso, cada resultado é um evento independente do outro.
Da mesma maneira, o nascimento de uma criança com um
determinado fenótipo é um evento independente em relação ao nascimento
de outros filhos do mesmo casal. Por exemplo, imagine uma casal que já
teve dois filhos homens; qual a probabilidade que uma terceira criança
seja do sexo feminino? Uma vez que a formação de cada filho é um evento
independente, a chance de nascer uma menina, supondo que homens e
mulheres nasçam com a mesma freqüência, é 1/2 ou 50%, como em qualquer
nascimento.
A regra do “e”
A teoria das probabilidades diz que
a probabilidade de dois ou mais eventos independentes ocorrerem
conjuntamente é igual ao produto das probabilidades de ocorrerem
separadamente. Esse princípio é conhecido popularmente como regra do “e”, pois corresponde a pergunta: qual a probabilidade de ocorrer um evento E outro, simultaneamente?
Suponha que você jogue uma moeda
duas vezes. Qual a probabilidade de obter duas “caras”, ou seja, “cara”
no primeiro lançamento e “cara” no segundo? A chance de ocorrer “cara”
na primeira jogada é, como já vimos, igual a ½; a chance de ocorrer
“cara” na segunda jogada também é igual a1/2. Assim a probabilidade
desses dois eventos ocorrer conjuntamente é 1/2 X 1/2 = 1/4.
No lançamento simultâneo de três dados, qual a
probabilidade de sortear “face 6” em todos? A chance de ocorrer “face 6”
em cada dado é igual a 1/6. Portanto a probabilidade de ocorrer “face
6” nos três dados é 1/6 X 1/6 X 1/6 = 1/216. Isso quer dizer que a
obtenção de três “faces 6” simultâneas se repetirá, em média, 1 a cada
216 jogadas.
|
 |
A regra do “ou”
Outro princípio de probabilidade diz que a ocorrência de dois eventos que se excluem mutuamente é igual à soma das probabilidades com que cada evento ocorre. Esse princípio é conhecido popularmente como regra do “ou”, pois corresponde à pergunta: qual é a probabilidade de ocorrer um evento OU outro?
Por exemplo, a probabilidade de
obter “cara” ou “coroa”, ao lançarmos uma moeda, é igual a 1, porque
representa a probabilidade de ocorrer “cara” somada à probabilidade de
ocorrer “coroa” (1/2 + 1/2 =1). Para calcular a probabilidade de obter
“face 1” ou “face 6” no lançamento de um dado, basta somar as
probabilidades de cada evento: 1/6 + 1/6 = 2/6.
Em certos casos precisamos aplicar tanto a
regra do “e” como a regra do “ou” em nossos cálculos de probabilidade.
Por exemplo, no lançamento de duas moedas, qual a probabilidade de se
obter “cara” em uma delas e “coroa” na outra? Para ocorrer “cara” na
primeira moeda E “coroa” na segunda, OU
“coroa” na primeira e “cara” na segunda. Assim nesse caso se aplica a
regra do “e” combinada a regra do “ou”. A probabilidade de ocorrer
“cara” E “coroa” (1/2 X 1/2 = 1/4) OU “coroa” e “cara” (1/2 X 1/2 = 1/4)
é igual a 1/2 (1/4 + 1/4).
|
 |
O mesmo raciocínio se aplica aos problemas da genética.
Por exemplo, qual a probabilidade de uma casal ter dois filhos, um do
sexo masculino e outro do sexo feminino? Como já vimos, a probabilidade
de uma criança ser do sexo masculino é ½ e de ser do sexo feminino
também é de ½. Há duas maneiras de uma casal ter um menino e uma menina:
o primeiro filho ser menino E o segundo filho ser menina (1/2 X 1/2 =
1/4) OU o primeiro ser menina e o segundo ser menino (1/2 X 1/2 = 1/4). A
probabilidade final é 1/4 + 1/4 = 2/4, ou 1/2.
Alelos múltiplos na determinação de um caráter
Como sabemos, genes alelos são os que atuam na determinação de um mesmo caráter e estão presentes nos mesmo loci (plural de lócus,
do latim, local) em cromossomos homólogos. Até agora, só estudamos
casos em que só existiam dois tipos de alelos para uma dada
característica (alelos simples), mas há caso em que mais de dois tipos
de alelos estão presentes na determinação de um determinado caráter na
população. Esse tipo de herança é conhecido como alelos múltiplos (ou
polialelia).
Apesar de poderem existir mais de
dois alelos para a determinação de um determinado caráter, um indivíduo
diplóide apresenta apenas um par de alelos para a determinação dessa
característica, isto é, um alelo em cada lócus do cromossomo que
constitui o par homólogo.
São bastante frequentes os casos de alelos múltiplos
tanto em animais como em vegetais, mas são clássicos os exemplos de
polialelia na determinação da cor da pelagem em coelhos e na
determinação dos grupos sanguíneos do sistema ABO em humanos.
Um exemplo bem interessante e
de fácil compreensão, é a determinação da pelagem em coelhos, onde
podemos observar a manifestação genética de uma série com quatro genes
alelos: o primeiro C, expressando a cor Aguti ou Selvagem; o segundo Cch, transmitindo a cor Chinchila; o terceiro Ch, representando a cor Himalaia; e o quarto alelo Ca, responsável pela cor Albina.
Sendo a relação de dominância → C > Cch > Ch > Ca O gene C é dominante sobre todos os outros três, o Cch dominante em relação ao himalaia e ao albino, porém recessivo perante o aguti, e assim sucessivamente. |
 |
O quadro abaixo representa as combinações entre os alelos e os fenótipos resultantes.
Genótipo
|
Fenótipo
|
| CC, C Cch, C Ch e C Ca |
Selvagem ou aguti
|
| CchCch, CchCh e CchCa |
Chinchila
|
| ChCh e Ch Ca |
Himalaia
|
| CaCa |
Albino
|
A diferença na cor da pelagem do coelho em relação à
cor da semente das ervilhas é que agora temos mais genes diferentes
atuando (4), em relação aos dois genes clássicos. No entanto, é
fundamental saber a 1ª lei de Mendel continua sendo obedecida, isto é,
para a determinação da cor da pelagem, o coelho terá dois dos quatro
genes. A novidade é que o número de genótipos e fenótipos é maior quando
comparado, por exemplo, com a cor da semente de ervilha.
O surgimento dos alelos múltiplos (polialelia)
deve-se a uma das propriedades do material genético, que é a de sofrer
mutações. Assim, acredita-se que a partir do gene C (aguti), por um erro acidental na duplicação do DNA, originou-se o gene Cch (chinchila).
A existência de alelos múltiplos é interessante para a espécie, pois
haverá maior variabilidade genética, possibilitando mais oportunidade
para adaptação ao ambiente (seleção natural).
Sistema ABO de grupos sanguíneos
A herança dos tipos sanguíneos do sistema ABO constitui um exemplo de alelos múltiplos na espécie humana.

Determinação dos grupos sanguíneos
utilizando soros anti-A e anti-B. Amostra 1- sangue tipo A. Amostra 2 -
sangue tipo B. Amostra 3 - sangue tipo AB. Amostra 4 - sangue tipo O.
|
A descoberta dos grupos sanguíneos
Por volta de 1900, o médico austríaco Karl
Landsteiner (1868 – 1943) verificou que, quando amostras de sangue de
determinadas pessoas eram misturadas, as hemácias se juntavam, formando
aglomerados semelhantes a coágulos. Landsteiner concluiu que
determinadas pessoas têm sangues incompatíveis, e, de fato, as pesquisas
posteriores revelaram a existência de diversos tipos sanguíneos, nos
diferentes indivíduos da população.
Quando, em uma transfusão, uma pessoa recebe um
tipo de sangue incompatível com o seu, as hemácias transferidas vão se
aglutinando assim que penetram na circulação, formando aglomerados
compactos que podem obstruir os capilares, prejudicando a circulação do
sangue.
Aglutinogênios e aglutininas
No sistema ABO existem quatro tipos de sangues: A, B, AB e O.
Esses tipos são caracterizados pela presença ou não de certas
substâncias na membrana das hemácias, os aglutinogênios, e pela presença
ou ausência de outras substâncias, as aglutininas, no plasma sanguíneo.
Existem dois tipos de aglutinogênio, A e B, e
dois tipos de aglutinina, anti-A e anti-B. Pessoas do grupo A possuem
aglutinogênio A, nas hemácias e aglutinina anti-B no plasma; as do grupo
B têm aglutinogênio B nas hemácias e aglutinina anti-A no plasma;
pessoas do grupo AB têm aglutinogênios A e B nas hemácias e nenhuma
aglutinina no plasma; e pessoas do gripo O não tem aglutinogênios na
hemácias, mas possuem as duas aglutininas, anti-A e anti-B, no plasma.
|
Veja na tabela abaixo a compatibilidade entre os diversos tipos de sangue:
ABO
|
Substâncias
|
%
|
Pode receber de
|
||||||||
Tipos
|
Aglutinogênio
|
Aglutinina
|
Frequência
|
A+
|
B+
|
A+
|
0+
|
A-
|
B-
|
AB-
|
O-
|
AB+
|
A e B
|
Não Contém
|
3%
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
A+
|
A
|
Anti-B
|
34%
|
X
|
X
|
X
|
X
|
||||
B+
|
B
|
Anti-A
|
9%
|
X
|
X
|
X
|
X
|
||||
O+
|
Não Contém
|
Anti-A e Anti-B
|
38%
|
X
|
X
|
||||||
AB-
|
Ae B
|
Não Contém
|
1%
|
X
|
X
|
X
|
X
|
||||
A-
|
A
|
Anti-B
|
6%
|
X
|
X
|
||||||
B-
|
B
|
Anti-A
|
2%
|
X
|
X
|
||||||
O-
|
Não Contém
|
Anti-A e Anti-B
|
7%
|
X
|
|||||||
Tipos possíveis de transfusão
As aglutinações que caracterizam as
incompatibilidades sanguíneas do sistema acontecem quando uma pessoa
possuidora de determinada aglutinina recebe sangue com o aglutinogênio
correspondente.
Indivíduos do grupo A não podem
doar sangue para indivíduos do grupo B, porque as hemácias A, ao
entrarem na corrente sanguínea do receptor B, são imediatamente
aglutinadas pelo anti-A nele presente. A recíproca é verdadeira:
indivíduos do grupo B não podem doar sangue para indivíduos do grupo A.
Tampouco indivíduos A, B ou AB podem doar sangue para indivíduos O, uma
vez que estes têm aglutininas anti-A e anti-B, que aglutinam as hemácias
portadoras de aglutinogênios A e B ou de ambos.
Assim, o aspecto realmente importante da
transfusão é o tipo de aglutinogênio da hemácia do doador e o tipo de
aglutinina do plasma do receptor. Indivíduos do tipo O podem doar sangue
para qualquer pessoa, porque não possuem aglutinogênios A e B em suas
hemácias. Indivíduos, AB, por outro lado, podem receber qualquer tipo de
sangue, porque não possuem aglutininas no plasma. Por isso, indivíduos
do grupo O são chamadas de doadores universais, enquanto os do tipo AB
são receptores universais.
|
 |
Como ocorre a Herança dos Grupos Sanguíneos no Sistema ABO?
A produção de aglutinogênios A e B são determinadas, respectivamente, pelos genes I A e I B. Um terceiro gene, chamado i, condiciona a não produção de aglutinogênios. Trata-se, portanto de um caso de alelos múltiplos. Entre os genes I A e I B há co-dominância (I A = I B), mas cada um deles domina o gene i (I A > i e I B> i).
Fenótipos
|
Genótipos
|
A
|
I AI A ou I Ai
|
B
|
I BI B ou I Bi
|
AB
|
I AI B
|
O
|
ii
|
A partir desses conhecimentos fica claro que se uma
pessoa do tipo sanguíneo A recebe sangue tipo B as hemácias contidas no
sangue doado seriam aglutinadas pelas aglutininas anti-B do receptor e
vice-versa.
O sistema MN de grupos sanguíneos
Dois outros antígenos forma encontrados na superfície das hemácias humanas, sendo denominados M e N. Analisando o sangue de diversas pessoas, verificou-se que em algumas existia apenas o antígeno M, em outras, somente o N e várias pessoas possuíam os dois antígenos. Foi possível concluir então, que existiam três grupos nesse sistema: M, N e MN.
|
Os genes que condicionam a produção desses antígenos são apenas dois: L M e L N (a letra L é a inicial do descobridor, Landsteiner). Trata-se de uma caso de herança medeliana simples. O genótipo L ML M, condiciona a produção do antígeno M, e L NL N, a do antígeno N. Entre L M e L N há co-dominância, de modo que pessoas com genótipo L ML N produzem os dois tipos de antígenos.
|
Transfusões no Sistema MN
A produção de anticorpos anti-M ou anti-N
ocorre somente após sensibilização (você verá isso no sistema RH).
Assim, não haverá reação de incompatibilidade se uma pessoa que pertence
ao grupo M, por exemplo, receber o sangue tipo N, a não ser que ela
esteja sensibilizada por transfusões anteriores.
O sistema RH de grupos sanguíneos
Um terceiro sistema de grupos sanguíneos foi
descoberto a partir dos experimentos desenvolvidos por Landsteiner e
Wiener, em 1940, com sangue de macaco do gênero Rhesus. Esses
pesquisadores verificaram que ao se injetar o sangue desse macaco em
cobaias, havia produção de anticorpos para combater as hemácias
introduzidas. Ao centrifugar o sangue das cobaias obteve-se o soro que
continha anticorpos anti-Rh e que poderia aglutinar as hemácias do
macaco Rhesus. As conclusões daí obtidas levariam a descoberta de um antígeno de membrana que foi denominado Rh (Rhesus), que existia nesta espécie e não em outras como as de cobaia e, portanto, estimulavam a produção anticorpos, denominados anti-Rh.
Há neste momento uma inferência evolutiva: se as
proteínas que existem nas hemácias de vários animais podem se assemelhar
isto pode ser um indício de evolução. Na espécie humana, por exemplo,
temos vários tipos de sistemas sanguíneos e que podem ser observados em
outras espécies principalmente de macacos superiores.
Analisando o sangue de muitos indivíduos da espécie
humana, Landsteiner verificou que, ao misturar gotas de sangue dos
indivíduos com o soro contendo anti-Rh, cerca de 85% dos indivíduos
apresentavam aglutinação (e pertenciam a raça branca) e 15% não
apresentavam. Definiu-se, assim, "o grupo sanguíneo Rh +” ( apresentavam o antígeno Rh), e "o grupo Rh -“ ( não apresentavam o antígeno Rh).
No plasma não ocorre naturalmente o anticorpo
anti-Rh, de modo semelhante ao que acontece no sistema Mn. O anticorpo,
no entanto, pode ser formado se uma pessoa do grupo Rh -, recebe sangue de uma pessoa do grupo Rh +.
Esse problema nas transfusões de sangue não são tão graves, a não ser
que as transfusões ocorram repetidas vezes, como também é o caso do
sistema MN.
A Herança do Sistema Rh
Três pares de genes estão envolvidos na herança do fator Rh, tratando-se portanto, de casos de alelos múltiplos.
| Para simplificar, no entanto, considera-se o envolvimento de apenas um desses pares na produção do fator Rh, motivo pelo qual passa a ser considerado um caso de herança mendeliana simples. O gene R, dominante, determina a presença do fator Rh, enquanto o gene r, recessivo, condiciona a ausência do referido fator. |
|
Doença hemolítica do recém-nascido ou eritroblastose fetal
Uma doença provocada pelo fator Rh é a
eritroblastose fetal ou doença hemolítica do recém-nascido,
caracterizada pela destruição das hemácias do feto ou do recém-nascido.
As conseqüências desta doença são graves, podendo levar a criança à
morte.
Durante a gestação ocorre passagem, através da
placenta, apenas de plasma da mãe para o filho e vice-versa devido à
chamada barreira hemato-placentária. Pode ocorrer, entretanto, acidentes
vasculares na placenta, o que permite a passagem de hemácias do feto
para a circulação materna. Nos casos em que o feto possui sangue fator
rh positivo os antígenos existentes em suas hemácias estimularão o
sistema imune materno a produzir anticorpos anti-Rh que ficarão no
plasma materno e podem, por serem da classe IgG, passar pela BHP
provocando lise nas hemácias fetais. A produção de anticorpos obedece a
uma cascata de eventos (ver imunidade humoral) e por isto a produção de
anticorpos é lenta e a quantidade pequena num primeiro. A partir da
segunda gestação, ou após a sensibilização por transfusão sanguínea, se o
filho é Rh + novamente, o organismo materno já conterá anticorpos para
aquele antígeno e o feto poderá desenvolver a DHPN ou eritroblastose
fetal.
O diagnóstico pode ser feito pela tipagem sanguínea
da mãe e do pai precocemente e durante a gestação o teste de Coombs que
utiliza anti-anticorpo humano pode detectar se esta havendo a produção
de anticorpos pela mãe e providências podem ser tomadas. Uma transfusão ,
recebendo sangue Rh -, pode ser feita até mesmo intra-útero já que
Goiânia está se tornando referência em fertilização in vitro. O sangue
Rh - não possui hemácias com fator Rh e não podem ser reconhecidas como
estranhas e destruídas pelos anticorpos recebidos da mãe. Após cerca de
120 dias, as hemácias serão substituídas por outras produzidas pelo
próprio indivíduo. O sangue novamente será do tipo Rh +, mas o feto já
não correrá mais perigo

Após o nascimento da criança toma-se medida profilática injetando, na mãe Rh-
, soro contendo anti Rh. A aplicação logo após o parto, destrói as
hemácias fetais que possam ter passado pela placenta no nascimento ou
antes. Evita-se , assim, a produção de anticorpos “zerando o placar de
contagem”. Cada vez que um concepto nascer e for Rh+ deve-se fazer nova
aplicação pois novos anticorpos serão formados.
Os sintomas no RN que podem ser observados são anemia (devida à destruição de hemácias pelos anticorpos), icterícia (a
destruição de hemácias aumentada levará a produção maior de bilirrubina
indireta que não pode ser convertida no fígado), e após sua
persistência o aparecimento de uma doença chamada Kernicterus que corresponde ao depósito de bilirrubina nos núcleos da base cerebrais o que gerará retardo no RN.
CITOLOGIA
Citologia
A fronteira das células
No mundo de hoje, é comum pensarmos em um país
como sendo uma porção de terra delimitada espacialmente das demais pela
presença de uma fronteira. Vamos pensar no caso do Brasil. Estamos
rodeados de mar em metade do nosso território e, na outra metade,
fazemos fronteira terrestre com outros nove países da America do Sul. Em
suas fronteiras, todos os países instalam uma alfândega, que é uma
repartição governamental de controle do movimento de entradas e saídas
das pessoas e de mercadorias para o exterior ou deles provenientes.
Com as células não é diferente. Cada uma delas
tem uma “área de fronteira”, representada pela membrana plasmática e,
nesta área, as células também possuem o seu “posto alfandegário”, as
proteínas. Assim como nas aduanas das fronteiras entre os países, essas
proteínas são as responsáveis pelo reconhecimento de substâncias vindas
de dentro ou de fora da célula como, por exemplo, hormônios.
O trabalho realizado por uma célula é semelhante
ao que acontece em uma fábrica, como a de televisores, por exemplo.
Através de portões, dá-se a entrada de diversos tipos de peças
destinadas as linhas de montagem. Para a fabricação e a montagem dos
aparelhos, são necessários energia e operários habilitados. É preciso,
ainda, um setor de embalagem para preparar a expedição do que é
produzido e uma diretoria para comandar todo o complexo fabril e manter o
relacionamento com o mundo externo. Tudo dentro dos limites
representados pelo muro da fábrica.

A célula possui setores semelhantes aos de uma
fábrica. Um limite celular, representado pela membrana plasmática,
separa o conteúdo da célula, o citoplasma, do meio externo. O
citoplasma, constituído por organóides e hialoplasma (ou citosol), um
material viscoso representa o setor produtivo. Um núcleo contendo o
material genético representa “a diretoria” da célula.
Os limites da célula viva
Uma célula viva é um compartimento microscópico,
isolado do ambiente por pelo menos uma barreira: a membrana plasmática.
Está é uma película extremamente fina e delicada, que exerce severa
“fiscalização” sobre todas as substâncias e partículas que entram e saem
da célula.

Dada a relativa fragilidade da membrana
plasmática, a maioria das células apresenta algum tipo de envoltório que
dá proteção e suporte físico à membrana. Entre esses envoltórios
destacam-se o glicocálix, presente na maioria das células animais, e a
parede celulósica, presente em células de plantas e de algumas algas.
Glicocálix
 |
Se isolássemos uma célula de nosso corpo, notaríamos que ela esta envolta por uma espécie de malha feita de moléculas de glicídios (carboidratos) frouxamente entrelaçadas. Esta malha protege a célula como uma vestimenta: trata-se do glicocálix (do grego glykys, doce, açúcar, e do latim calyx, casca envoltório).
|
Diversas funções têm sido sugeridas para o glicocálix. Acredita-se que, além de ser uma proteção
contra agressões físicas e químicas do ambiente externo, ele funcione
como uma malha de retenção de nutrientes e enzimas, mantendo um microambiente adequado ao redor de cada célula. Confere às células a capacidade de se reconhecerem,
uma vez que células diferentes têm glicocálix formado por glicídios
diferentes e células iguais têm glicocálix formado por glicídios iguais.

Parede celulósica
As células vegetais possuem um envoltório externo, espesso e relativamente rígido: a parede celulósica, também chamada membrana esquelética celulósica;
Paredes celulósicas primárias e secundária
Células vegetais jovens apresentam uma parede celulósica fina e flexível, denominada parede primária.

A parede primária é elástica, de modo a permitir o
crescimento celular. Depois que a célula cresceu e atingiu o tamanho e a
forma definitivos, forma-se a parede secundária, mais
espessa e rígida. A celulose que constitui a parede secundária é
secretada através da membrana plasmática, e se deposita entre esta e a
superfície interna da parede primária, na qual adere fortemente.

Constituição da parede celulósica
A parede das células vegetais é constituída por
longas e resistentes microfibrilas do polissacarídeo celulose. As
microfibrilas celulósicas se mantém unidas por meio de uma matriz
formada por glicoproteínas (proteínas ligadas a açucares), hemicelulose e
pectina (polissacarídeos).

A estrutura molecular da parede celulósica aplica
o mesmo princípio do concreto armado, no qual longas e resistentes
varetas de ferro estão mergulhadas em uma argamassa de cimento e pedras.
Na parede celular, as microfibrilas de celulose
correspondem às varetas de ferro do concreto, enquanto as glicoproteínas
e os polissacarídeos da matriz correspondem à argamassa.
domingo, 5 de agosto de 2012
terça-feira, 22 de maio de 2012



sábado, 5 de maio de 2012
terça-feira, 17 de abril de 2012
Vírus

VÍRUS
Características Gerais e Classificação
Introdução:
Se bem que doenças de animais e plantas, causadas por vírus, sejam tão velhasquanto a existência do homem sobre a face da terra, seu reconhecimento etilógico
só começou a ser feito no último decênio do século passado, quando Iwanowski,
trabalhando em Mosaico do Fumo, provou que suco de plantas doentes permanecia
infectivo depois da passagem pelo filtro bacteriológico. Essa descoberta foi logo
seguida por caraterização idêntica de várias doenças como a febre aftosa, a
poliomielite e a varíola. Até por volta de 1920, entretanto, devido ao fato de sua
diferenciação com microrganismos patogênicos residir tão somente em sua
filtrabilidade através do filtro bacteriológico e sua invisibilidade ao microscópio
composto, foi dado ênfase em estudos clínicos, patológicos e epidemiológicos
dessas doenças que, apesar de serem, obviamente, ocasionadas por agentes infeciosos transmissíveis, não podiam ser atribuídas a microrganismos visíveis.
A partir de então, estudos bioquímicos, sorológicos, eletromicroscópicos, etc.,
permitiram compreender melhor a natureza do vírus, um importante grupo de
agentes produtores de doenças, responsável por grandes problemas patológicos
no homem nos animais e nas plantas.
Características Filtrabilidade:
Essa característica, que serviu de base para a descoberta do vírus, originou adenominação vulgar de "vírus filtrável", termo que teve de ser abandonado pelo
fato de, posteriormente, se descobrir que existem bactérias filtráveis, como certos
espiroquetas, e vírus não filtráveis, como o vírus do Mosaico Comum do feijoeiro e
o do "Crinkle Mosaic" da batatinha. A filtrabilidade do vírus não é uma pura
conseqüência de seu diminuto tamanho. Tipo de filtro, temperatura, ph, carga
elétrica do vírus e do filtro, quantidade de pressão exercida sobre o filtro, natureza
do fluído da suspensão e duração da filtração, são fatores que devem ser levados
em consideração ao se determinar a filtrabilidade do vírus. Atualmente se dispõe de
filtros de colódio, as Membranas de Gradocol, em que o tamanho dos poros é muito
mais importante do que nos filtros bacteriológicos de porcelana.
Natureza Corpuscular
Desde 1898 se suspeita da natureza corpuscular do vírus, com a teoria do"contagium vivum fluidum", denominação que Beijerink emprestou de Fracastorius
(1546) para caracterizar o filtrado infectivo do suco de fumo com mosaico. A guisa
de esclarecimento, contagiam era uma substância derivada do corpo do doente e
que, passando de um indivíduo para outro, transmitia a doença e a teoria do
contagiam vivum foi criada por Fracastorius, quando postulou a idéia de que o
contagiam fosse devido a agentes vivos (seminaria). Entretanto, por muitos anos,
essa característica foi um ponto altamente controvertido pois a ciência ainda não
estava preparada para comprová-la. Apesar disso, mesmo antes da descoberta de
microscópio eletrônico, em 1938, Wendell Stanley (1935) já mostrava evidências
irrefutáveis sobre a natureza corpuscular do vírus ao cristalizar o vírus do Mosaico
do Fumo. Hoje, acostumamos com representações esquemáticas e
eletromicrográficos dos vírus, dificilmente imaginamos que essa característica
tivesse sido um pomo de discórdia no passado.
Dimensões do vírus
As dimensões dos vírus, evidenciadas por estudos eletromicroscópicos, deultrafiltração e ultracentrifugação, variam de 10 a 350 milimicra de diâmetro; o
comprimento chega até 2.000 milindrica (vírus da Tristeza do Citrus). A guisa de
comparação, os glóbulos vermelhos do sangue humano têm 7.500 milimicra de
diâmetro e, dentro de uma célula bacteriana, podem caber mais de 1 milhão de
partículas de vírus.
Morfologia, composição e estrutura
O microscópio eletrônico permite visualizar diretamente as característicasmorfológicas dos vírus que podem ser: esféricos, como os vírus da influenza e da
encefalite japonesa; cilíndricos, com a maioria dos vírus de plantas; cúbicas, como
o vírus da vaccinia; e espermatozoidal, (imagem)como os bacteriófagos.
A organização estrutural das partículas de vírus, evidenciada pela técnica da

configurações estridimensionais que admitem os seguintes tipos de simetria:
cúbica, helicoidal e binária.
Um vírus é composto de ácido nucleico ( DNA ou RNA) e uma parede de proteína.
Os ácidos nucleicos se localizam no interior das proteínas. As envoltório protéico
se dá o nome de capsídeo, o qual, por sua vez, é formado pelos capsômeros,
unidades estruturais simétricas dispostas em ordem determinada. O capasídeo
mais o ácido nucleico é o nucleocapsídeo. Virion é a partícula madura do vírus e
pode ser sinônimo de nucleocapsídeo ou nucleocapsídeo dentro de um invólucro.
Invólucro é uma membrana de origem celular que pode envolver o capsídeo. O
termo vírus se aplica num sentido mais amplo, incluindo os diferentes estágios do
seu desenvolvimento.
O material genético DNA ou RNA. nunca se encontram simultaneamente no mesmo
vírus, o que constitui uma característica singular quando comparados com as
células de outros organismos vivos. O DNA é o portador de informação genética em
todos os organismos vivos. Em alguns vírus, o material genético é o DNA e, em
outros, o RNA. Os vírus diferem no seu conteúdo de DNA ou RNA. Os vírus de
plantas contêm RNA, exclusivamente; os vírus de animais, DNA ou RNA e os vírus
de bactérias (bacteriófagos) DNA, comumente. Os vírus mais complexos contêm,
além das nucleoproteínas, outros compostos como lipídeos, carboidratos,
vitaminas e vestígios de metais.
Transmissibilidade
A transmissibilidade é uma característica fundamental dos vírus, como também o épara outros agentes causadores de doenças. É interessante citar que, já em 1901,
nos primórdios da Virologia, se chegava à conclusão de que a Febre Amarela não
era transmitida pelo contato físico de pessoas doentes e sadias havendo
necessidade do concurso de um mosquito vetor na cadeia de transmissão de
homem a homem. Por muitos anos, a transmissibilidade do vírus foi a única
 evidência experimental de sua existência como uma entidade independente. Os
evidência experimental de sua existência como uma entidade independente. Osconhecimentos sobre a transmissibilidade são essenciais para se tentar prevenir ou
circunscrever a transmissão natural.
O vírus de plantas são transmitidos, na natureza, principalmente por meio de
enxertia, insetos vetores e contato membrânico. Existem casos de transmissão pelo
grão de pólen por fungos do solo, por nematóides e por ácaros, mas são raros.
Entretanto, para fins de considerações que os vírus podem ser disseminados a
longas distâncias por meio de sementes (Mosaico Comum do Feijoeiro), Mosaico da
Alface, TWV em tomateiro e órgãos de propagação vegetativa (mosaico e
raquitismo da cana-de-açúcar, enrolamento da batatinha, etc.)
A transmissão por enxertia é o método mais universalmente aplicável, requerendo
somente que o vírus se torne sistêmico. Assim se transmitem, nas condições
naturais, por exemplo, os vírus da Tristeza, Sacarose, Exocorte e Xiloporose do
Citrus. Experimentalmente, vírus comum a duas espécies vegetais que não se
enxertam podem ser transmitidos por enxerto de Cuscuta sp., fanerógamo parasita.
A transmissão por contato mecânico, muito usado em estudos de inoculação
artificial, é um método muito importante para vírus como o TWV e o Mosaico das
Cucurbitáceas que pelo simples contato do lavrador de uma planta doente para a
sadia pode transmitir o vírus.
A transmissão por insetos vetores é o método mais comum na natureza. Dentre os
insetos vetores os Afídeos constituem o grupo mais numeroso, sendo responsável
pela transmissão de mais ou menos 90 vírus diferentes. Dentre os afídeos sabe-se
que o Myzus persicae é transmissor de mais de 50 vírus diferentes. Há
determinados vírus que são transmitidos por vários insetos, como por exemplo o
vírus do mosaico do pepino que é transmitido por algumas dezenas de insetos.
Também existem vírus transmissíveis por apenas um inseto, como no caso do vírus
da beterraba açucareira transmitida por Circulifer tenellus. Os trips se caracterizam
por transmitir somente uma espécie de vírus de planta, como, por exemplo, no caso
de Frankliniella paucispinosa que transmite somente o vírus do vira-cabeça.
A transmissão do vírus pelos insetos pode Ter um caráter persistentes ou não
persistentes. Diz-se que o vírus é persistente quando, após um longo período
mínimo de alimentação em plantas doentes, para aquisição e após um longo
período de retenção no inseto, geralmente por toda a vida. Por exemplo, vírus do
enrolamento das folhas da batatinha se multiplica com maior eficiência no inseto
vetor Myzus persicae com maior período de alimentação; apresenta um período
latente de 24 horas e, o pulgão guarda o vírus não persistentes são adquiridos pelo
vetor num curto período de alimentação (10 horas no máximo) e são retidos por um
período máximo de 24 horas. Por exemplo, o vírus do mosaico da beterraba é
adquirido por Myzus persicae em 2 horas e é retido por apenas 3 horas.
Parasitismo celular obrigatório
A mais importante característica dos vírus é o parasitismo celular obrigatório. Sebem que partículas individuais dos vírus sobrevivam por períodos variáveis em
ambientes extracelulares, a sobrevivência da espécie depende inteiramente de
multiplicação intracelular e, portanto, de sucessivos ciclos de associação com
organismos hospedeiros mais evoluídos (bactérias, plantas e metazoários). Com
efeito, todas a tentativas de cultivá-los em meio de cultura inanimado fracassaram.
Dessa características resulta toda a importância dos vírus pois, frequentemente, ao
parasitismo obrigatório se associa a patogênese.
Do parasitismo obrigatório decorrem ainda duas importantes consequências: a
formação de inclusões intracelulares e o desenvolvimento de técnicas especiais de
cultura artificial do vírus. Inclusões intracelulares (protoplásmicas e ou
intranucleares) já são conhecidas em várias viroses, tanto vegetais como animais.
Tais inclusões são interpretadas, atualmente, como colônias intracelulares do vírus.
Ex. inclusões cristalinas hexagonais como em TWV, inclusões em massa compacta
como em Tristeza do Citrus, inclusões cilíndrico-lamelares como no Mosaico
Comum do feijoeiro, inclusões nucleares como no vírus da Gomphrena, etc.
Em todas as técnicas especiais de cultura artificial dos vírus é imprescindível a
presença de células vivas. Os métodos mais usados para o cultivo de vírus de
interesse médico são a cultura de tecido e a cultura do ovo embrionado. O vírus de
interesse fitopatológico não são muito estudados sobre esse aspecto.
Infecção e multiplicação
Os vírus se aderem à parede celular, penetram, se multiplicam, amadurecem e,finalmente, se liberam. Nos vírus de plantas, aparentemente, não existe a
capacidade de penetração, havendo necessidade de colocar a partícula do vírus em
contato direto com o ambiente intracelular.
Nos vírus de plantas, a primeira fase do processo de infecção é a adsorsão. O vírus
deve ser introduzido na célula, injuriada mas não morta, entrando em contato
íntimo e se unindo com a substância viva da célula. Essa união é instantânea de
modo que a lavagem imediata do tecido inoculado com água não reduz a infecção.
Comprovou-se, experimentalmente, que a imersão de folhas de Nicotiana glutinosa,
logo após inoculação como RNA do TWV, em uma solução de RNA se, suficiente
para destruir o RNA não reduz o número de lesões locais.
Aparentemente, os vírus de plantas, logo após a adsorsão, de desfazem da parte
 protéica liberando o RNA. Segue-se um período de latência ou de eclipse durante o
protéica liberando o RNA. Segue-se um período de latência ou de eclipse durante oqual o vírus não pode ser detectado. A duração desse período depende da
temperatura, do conteúdo de vírus no inoculo e da sensibilidade do método. Depois
dessa fase o vírus se torna detectável e o seu conteúdo nos extratos sucessivos
aumenta rapidamente.
O fenômeno dominante da infecção por vírus é a replicação intracelular. Sabe-se,
hoje, que os vírus de plantas consistem de dois componentes químicos, RNA
infeccioso e a proteína do vírus. Essa proteína não exibe nenhuma atividade
enzimática conhecida, não podendo iniciar por si a infeção e, aparentemente,
servindo somente como capa protetora para o RNA. (nos bacteriófagos a capa
protéica exibe atividade enzimática e a penetração assume um caráter ativo). O
RNA, por seu lado, possui a capacidade de causar infeção que resulta na formação
de partículas típicas do vírus. Parece, portanto, carregar a informação genética para
reproduzir não somente a si próprio mas também a parte protéica do vírus.
Especificidade de Hospedeiros e Tecidos
Os vírus, tanto de plantas como de animais, apresentam uma gama determinada dehospedeiros. Assim, o vírus da febre amarela urbana tem como hospedeiros
somente o homem (transmissor: mosquito do gênero Aedes); o da febre amarela
silvestre, o macaco e o homem (transmissor Haemogogus); o da Tristeza do Citrus,
somente plantas cítricas; TWV pelo menos 74 espécies vegetais distribuídas em 14
famílias. Em vírus animais e especificidade vai até o nível histológico, servindo de
base para classifica-los em vírus: vírus dermotrópicos (varíola, varicela, sarampo,
rubéola, etc.), vírus pneumotrópicos (gripe, resfriado, etc.) vírus neurotrópicos
(raiva, poliomielite, encefalites, etc.), vírus hepatotrópicos (febre-amarela, hepatite)
e vírus linfo e glandulotrópicos (caxumba, linfogranuloma inguinal).
NATUREZA ANTIGÊNICA
Muito antes da descoberta dos vírus, já se sabia que doenças hoje conhecidas eramcausadas por vírus, como por exemplo a varíola, conferiam resistência contra
incidências subsequentes. A vacina contra a varíola se baseia, ainda hoje, na
descobertas de Jenner (1798) de que o vírus do "cow-pox"(varíola bovina) imuniza
contra o "small-pox"(varíola humana).
Proteínas introduzidas no corpo animal, por via parenteral, sendo elas estranhas ao
corpo do animal, induz a formação de substância que reagem especificamente com
as proteínas injetadas. Estas proteínas estranhas constituem os antígenos e as
substâncias induzidas, os anticorpos. Sendo os vírus de natureza nucleoproteica
tem essa propriedade antigêno que serve de base para os métodos sorológicos
usados em Virologia. Especula-se, atualmente, se as plantas possuem essa
capacidade de formação de anticorpos, comprova somente em animais. Estirpes
fracas do vírus da tristeza dos citros conferem resistência às estirpes mais severas
do mesmo vírus. A natureza desse fenômeno, entretanto, não esta esclarecida.
VARIABILIDADE
A variabilidade em vírus é reconhecida há muito tempo mas só recentemente temsido possível apreciar a sua extensão e especular as possíveis causas. Essa
variabilidade foi inicialmente observada em relação à patogenicidade e gama de
hospedeiros mas, atualmente, se conhecem evidência de variabilidade em certos
vírus para quase todas as características examinadas: constituição de
aminoácidos, capacidade de forma particular nucleoproteína típica, capacidade de
ser transmitida por determinado vetor, forma cristalina, resistência a tratamento
inativantes, etc. Evidentemente, variações em patogenicidade são as que
despertam maior interesse. Por exemplo, acontece, frequentemente, com vírus de
animais que, sendo inoculados em série no organismo de certos hospedeiros ou
após repetidas subculturas em meio artificial, modificam pouco a pouco a sua
virulência original até certo limite que se mantém estável. E essa perda de
virulência é irreversível. Por exemplo, o vírus da raiva colhido diretamente do cão
(vírus da ruas), passado varias vezes em coelhos, transforma-se no chamado vírus
fixo. Desse fenômeno se tira excelente proveito no preparo de vacinas.
CONCEITO DE VÍRUS
Vírus são partículas infecciosas, de natureza nucleoproteica, de dimensõesgeralmente inferiores a 0,2 micra e, consequentemente, geralmente filtráveis
em filtros bacteriológicos e visíveis somente ao microscópio eletrônico. São
parasitas intracelulares obrigatórios, formando geralmente só em presença
de células vivas e dão facilmente lugar a mutações. Induz a célula parasita a
formar réplicas, tanto do ácido nucleico como da capa protéica.
CLASSIFICAÇÃO E NOMENCLATURA
A classificação e a nomenclatura de vírus é, ainda hoje, um ponto controvertido.Inicialmente, os vírus foram denominados de acordo com o nome da doença que
ocasionavam e, apesar de muitas tentativas de introduzir novas nomenclaturas
científicas, é, ainda, a mais universalmente adotada entre os fitopatologistas. Assim
se conhecem, por exemplo, o vírus do mosaico do fumo (VMF ou TMV), o vírus da
vira cabeça do tomateiro, o vírus da tristeza do Citrus, o vírus do mosaico comum
do feijoeiro, etc. Evidentemente, tal nomenclatura foge à regra geral de
denominação de outros agentes causadores de doenças e pode-se considerá-la
comum ou vulgar (nome comum do citros). Como se pode observar, essas
denominações comuns se baseiam principalmente em sintomatologia das doenças
ocasionadas e sendo a sintomatologia um caráter variável, de acordo com o
ambiente e com hospedeiro, levou a muitas confusões, um mesmo vírus (como o
TMV e o vírus do mosaico das cucurbitáceas) sendo identificada várias vezes como

Johnson (1927), observando que sintomas, quando apropriadamente interpretadas
em estudos comparativos, tinham características diagnosticas de algum valor para
classificar os vírus do fumo mas que era difícil dar um nome descritivo para todos
os vírus que ocorrem num determinado hospedeiro, sugeriu uma nomenclatura
baseada em hospedeiro e prioridade de constatação. No seu sistema o vírus do
mosaico do fumo (VMF) se denominaria Tabaco vírus 1 e os outros vírus do fumo
receberia um número em ordem de sua descoberta, Tal sistema não tem,
atualmente, nenhum valor, pois um número nada caracteriza e o grande número de
vírus de um determinado hospedeiro dificulta a associação com características
importantes dos vírus.
Smith (1937) propôs a latinização do sistema de Johnson e , assim, o VMF se
denominaria Nicotiana vírus 1 , mantendo-se, ainda, as mesmas desvantagens do
sistema de Johnson.
Bennett (1939) sugeriu a substituição dos números por um termo que
caracterizasse uma propriedade importante do vírus e o VMF se chamaria Tobacco
vírus altathermus ou Nicotiana vírus altathermus (elevado ponto térmico de
inativação). Se em investigações subsequentes se provasse que o vírus fossem
organismos vivos, adotar-se-ia a denominação binomial Paracrystalis altathermus ;
se, pelo contrário, se provasse serem os vírus compostos químicos, o VMF, por
exemplo, se denominaria Altathermovir.
Holmes (1939) sugeriu a nomenclatura binomial - trinomial latinizada pela qual o
VMF se denominaria Marmor tabaci e suas linhagens M. tabaci var. vulgare, M.
tabaci var. aucuba, M. tabaci var. deformans, etc. Holmes propunha ainda a criação
do reino Vira , incluindo todos os vírus, com duas divisões: Zoophagi , para os
vírus de animais e Phytophagin , para os vírus de plantas.
Os Phytophagi foram subdivididos em duas classes: Schyzophyto phagi
(bactériofagos) Spermatophytophagi (vírus de plantas superiores). Estes admitiam
as seguintes famílias e gêneros: Chlorogenaceae (gen. Chlorogenus), Marmoraceae
(gen. Marmor), Annilaceae (gen. Annulus), Gallaceae (gen. Galla), Acrogenaceae
(gen. Acrogenus), Rugaceae (gen. Ruga), Coriaceae (gen. Corium), Nanaceae(gen.
Nanus), Savoiaceae (gen. Savoia) e Lethaceae (gen. Lethum). Segundo Holmes, tal
sistema de classificação tem as vantagens de agrupar os vírus de acordo com
similaridades fundamentais como testes sorológicos, e imunológicos e tipos de
doença.
Fawcett (1940), propôs a nomenclatura binomial em que o nome genérico era obtido
do hospedeiro mais o sufixo vir e o nome específico de alguma característica da
virose ou do vírus.
Assim, o vírus da sacarose do Citrus foi denominado de Citrivir psorosis, o vírus do
vira-cabeça da beterraba Betavir eutetticola etc.
Além dessas nomenclaturas foram sugeridas muitas outras mas nenhuma
conseguiu aceitação geral, estando, ainda hoje, a taxonomia de vírus num
verdadeiro cáos. Entretanto, já há um esforço, em âmbito internacional, visando
padronizar a nomenclatura e a classificação dos vírus. Nesse sentido, o Comitê
Provisório de Nomenclatura de vírus (P.C.N.V.) da Associação Internacional das
Sociedades Microbiológicas (1965) recomendou a adoção provisória do sistema de
classificação de Lwoff, Horne e Tournier (L.H.T.), por ser, no momento,
aparentemente, o mais adequado.
O sistema L.H.T., caracteriza os vírus como entidades que exibem durante o seu
"ciclo vital" uma partícula infecciosa contendo apenas um tipo de ácido nucleico.
Baseia-se, principalmente, em natureza do ácido nucleico, morfologia, estrutura e
simetria das partículas de vírus. Primeiramente, os vírus são em dois grupos: D-
(deoxyvira), contendo DNA e R- (ribovira), contendo RNA. Esses grupos são
subdivididos, de acôrdo com sua simetria, em H- com simetria helicoidal (classes
Deoxyhelica e ribohelica). C- com simetria cúbica (classe Deoxybinala). Todos os
virions pertencem a uma das duas categorias: N- capsídeo sem envólucro e,
capsídeo com envólucro. Os grupos são posteriormente subdivididos, de acordo
com o número de capsômeros, para os virions RCN, 32c. O quadro abaixo mostra o
sistema L.H.T. sugerindo pelo P.C.N.V.
A nomenclatura proposta por PCNV baseia-se nomes latinos ou gregos-latinizados,
escolhendo-se uma espécie típica para cada gênero. A espécie típica dá o nome à
família. Os nomes genéricos terminam em vírus. Exp. Protovirus tabaci (TMV).
Os nomes das famílias, derivadas dos nomes genéricos, terminam em VIRIDAE.
EXP. FAM. Protoviridae (T.M.V.).
Vídeo:
Assinar:
Comentários (Atom)

-
 Conceitos sobre circulação Circulação aberta: tipo de circulação em que o sangue ou hemolinfa sai do interior dos vasos e entra em conta...
Conceitos sobre circulação Circulação aberta: tipo de circulação em que o sangue ou hemolinfa sai do interior dos vasos e entra em conta... -
 O Lixo (RESÍDUOS SÓLIDOS) A natureza não é algo que está simplesmente fora do homem. O que seria dos nossos pulmões se faltas...
O Lixo (RESÍDUOS SÓLIDOS) A natureza não é algo que está simplesmente fora do homem. O que seria dos nossos pulmões se faltas...